
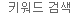


- 고객지원
- 업무안내│제품문의
- 전화번호│02-2081-2510
- Email │support@takara.co.kr
- 대전지사
- 업무안내│대전/충청지역 주문, 제품문의
- 전화번호│042-828-6525
- Email │tkbd@takara.co.kr
- 업무시간안내
- [ 평 일 ] 09 : 00 ~ 18 : 00 │ [ 점심시간 ] 12 : 00 ~ 13 : 00
- 토·일요일, 공휴일은 휴무입니다.
[Cancer Research] HLA typing


Cellartis human iPS cell-derived beta cells for diabetes drug discovery and disease modeling
다카라바이오는 당뇨병 및 대사 증후군 연구 모델과 신약 개발을 위한 hiPSC 유래 beta cell의 적용성을 확인하였다.
Introduction
당뇨나 다른 대사 증후군의 연구자에게 정확하고 재현 가능한 질병 모델의 제작은 어려움으로 남아 있다. 현재까지는 인슐린을 생산하는 beta cell을 포함하는 호르몬 생산 세포 군집인 췌장의 랑게르한스섬 (primary pancreatic islets of Langerhans)을 gold standard로 사용되고 있다. 다만 이 Primary cell은 샘플을 확보하기가 어렵고 비용이 많이 소요될 뿐 아니라, 유래한 donor의 유전적 차이로 인해 실험 결과에 변동이 생길 수 있다. 이를 대체하기 위해 단일 기증자에서 유래한 hiPSC로부터 분화한 beta cell이 신약 개발을 위한 효과적인 스크리닝과 insulin 생산 및 분비에 대한 생리학적 실험 모델로 사용될 수 있다.
일관적이고 신뢰도 높은 Beta cell 공급을 위해, 다카라바이오는 췌장 발달 과정을 모방하는 배양 조건을 최적화한 4 step의 표준 분화 프로토콜을 개발하였다 (그림 1). hiPSC는 먼저 내배엽 세포 (definitive endoderm cell)로 분화되고, 이후 pancreatic endoderm, endocrine progenitor cell 단계를 거쳐 최종적으로 성숙한 beta cell로 분화가 완료된다.
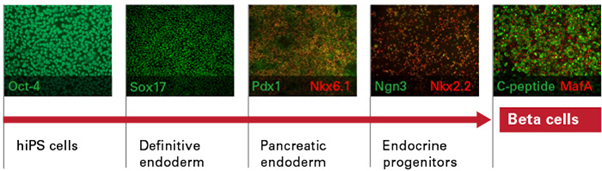
그림 1. hiPSC의 beta cell 분화 과정
Our differentiation strategy is based on a four-step protocol that resembles normal embryonic development of beta cells. hiPS cells are first differentiated into Sox17-positive definitive endoderm. Following endodermal commitment, the cells are further differentiated into Pdx1/Nkx6.1 double-positive pancreatic endoderm. Next, differentiation continues into endocrine progenitor cells, which are Ngn3/Nkx2.2 double-positive. Finally, the progenitors mature into terminally differentiated beta cells that are double positive for C-peptide and MafA. Abbreviations: Sox17 = SRY (sex determining region Y)-box 17; Pdx1 = pancreatic and duodenal homeobox 1; Nkx6.1 = NK6 homeobox 1; Ngn3 = Neurogenin 3; Nkx2.2 = NK2 homeobox 2; C-peptide = Connecting peptide; and MafA = MAF BZIP Transcription Factor A.
다카라바이오는 서로 다른 Donor로부터 얻은 hiPSC (ChiPSC12, ChiPSC22)로부터 분화된 총 두 종의 hiPSC 유래 beta cell을 판매하고 있으며, 제품은 세포와 coating substrate, basal media, supplement를 포함한 all-in-one 형태로 제공된다.
지속적인 생산이 가능한 Cellartis® beta cell은 성숙한 beta cell에서 나타나는 특징을 가질 뿐만 아니라, lot에 따른 차이를 최소화 하였고, 개인 간의 차이를 반영하는 기능 평가가 가능하기에 실험에 높은 신뢰성을 부여한다. 당뇨병 연구자들은 Donor HLA type에 따른 변이를 연구할 수 있으며, 역학 연구에 따르면 HLA-A*02:01을 포함하는 class Ⅰ 대립 유전자가 1형 당뇨병에 대한 감수성을 증가시킬 수 있다고 알려져 있다 (Marron et al. 2002). C12와 C22의 HLA type은 표 1에서 확인할 수 있다.
표 1. ChiPSC12와 ChiPSC22의 donor 및 HLA type 정보
The beta cells originate from donors with different HLA types. ChiPSC12 was sourced from skin fibroblasts from a healthy 24-year-old European/North African male volunteer (76 kg/177 cm). ChiPSC22 was sourced from skin fibroblasts from a healthy 32-year-old European/North African male volunteer (74 kg/179 cm) with a diabetes-susceptible HLA type, HLA-A*02:01.
hiPS cell-derived beta cells express beta-cell-specific genes
다카라바이오는 성숙한 Beta cell에서 존재하는 것으로 알려진 mRNA 전사체가 hiPSC 유래 beta cell에서 발현하는 지 먼저 분석하였다. hiPSC 유래 beta cell에서 모든 전사체가 확인되었으며, 발현 수준을 primary 랑게르한스섬 (islet of Langerhans)과 비교하였다 (그림 2). 특히, 각 Donor의 hiPSC 유래 beta cell의 모든 lot에서 유사한 수준의 mRNA 발현을 확인하였고, lot 간의 높은 재현성을 입증하였다.
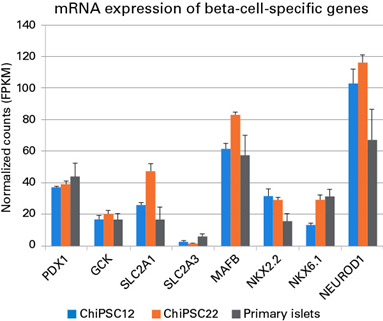
그림 2. hiPSC 유래 beta cell에서의 주요 mRNA 발현
mRNA expression analysis, as quantified by RNA-seq, was performed on Cellartis beta cells from C12 and C22 on Day 14 post-thawing (n = 3 batches per cell line) and on primary islets (n = 5 donors). Data are presented as the mean values ± SEM.
*PDX1 = pancreatic and duodenal homeobox 1; GCK = glucokinase; SLC2A1, SLC2A3 = Solute Carrier Family 2 Member 1 and 3; MAFB = MAF BZIP Transcription Factor B; NKX2.2 = NK2 homeobox 2; NKX6.1 = NK6 homeobox 1; and NEUROD1 = Neurogenic differentiation 1.
hiPS cell-derived beta cells synthesize insulin
Beta cell의 주요 기능은 insulin을 합성하고 분비하는 것으로, insulin 합성 과정을 살펴보면 proinsulin이 C-peptide 분자 하나와 insulin 분자 하나, 즉 1:1로 나뉘어 진다 (Ashby & Frier 1981; Horwitz et al. 1975). 따라서, C-peptide와 insulin의 동시 발현이 insulin 생산 능력의 평가 지표로 평가될 수 있다. 그림 3에서 C12와 C22 유래 beta cell 모두에서 C-peptide와 인슐린의 발현이 높은 상관관계를 보임으로써, 세포가 insulin 생산 능력을 갖추었다는 것을 확인할 수 있다.
그림 3. hiPSC 유래 beta cell에서의 insulin 합성
Staining for C-peptide and insulin was performed by immunofluorescence on Cellartis beta cells at Day 14 post-thawing. Colocalization of insulin and C-peptide indicates intracellular insulin synthesis. Scale bar = 100 ㎛.
hiPS cell-derived beta cells express key markers of mature beta cells
몇몇 전사 인자들은 Beta cell의 발달, 증식, 기능에 중요한 역할을 하며, 성숙한 beta cell은 Pdx1, Nkx6.1, 인슐린을 발현한다. Pdx1은 beta cell의 수명을 조절하는 역할로, beta cell이 발현하는 유전자를 활성화 하고 alpha cell에서 발현하는 유전자를 억제한다 (Gao et al. 2014). 반면 Nkx6.1이 결핍된 세포의 경우 delta cell의 분자적 특성을 보인다 (Taylor, Liu, and Sander 2013).
MafA는 성숙한 beta cell marker로써, 분화 과정에서는 발현되지 않아 다른 전사 인자와 확연히 구분된다. 또한 Insulin과 Nkx6.1이 동시 발현하는 것으로 성숙한 hiPSC 유래 beta cell를 확인할 수 있다 (Ma et al. 2018; Pagliuca et al. 2014; Rezania et al. 2014). 그림 4에서는 Cellartis® beta cell에서 insulin과 C-peptide와 함께 주요 beta cell marker인 Nkx6.1, Pdx1, MafA가 발현됨을 확인할 수 있다.
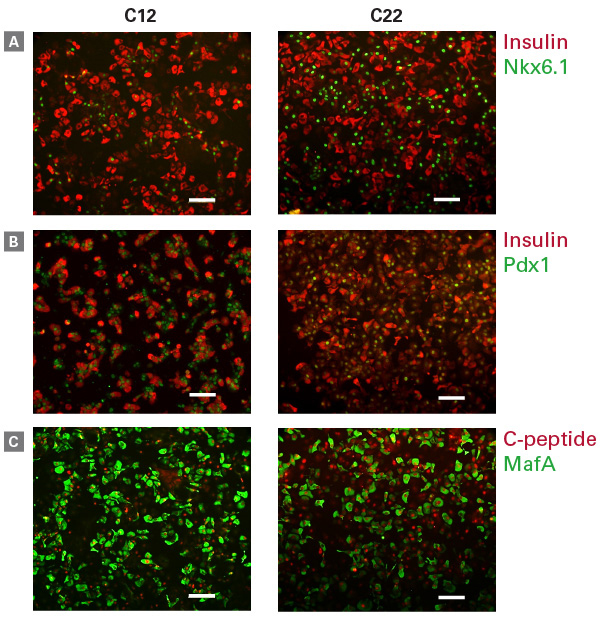
그림 4. hiPSC 유래 beta cell에서 확인한 key beta cell protein의 동시 발현
Staining for insulin and Nkx6.1 (Panel A), insulin and Pdx1 (Panel B), and C-peptide and MafA (Panel C) was performed on beta cells on Day 14 post-thawing. Scale bar = 100 ㎛.
뿐만 아니라, flow cytometry을 이용하여 insulin과 Nkx6.1를 동시 발현하는 세포의 정량 분석 결과를 그림 5에서 확인할 수 있다. 특히 insulin과 Nkx6.1을 동시에 발현하는 세포의 경우, C12과 C22에서 그 비율이 다르게 나타났으며, 이를 통해 HLA type에 따라 일부 유전자 발현에 차이를 확인할 수 있다.
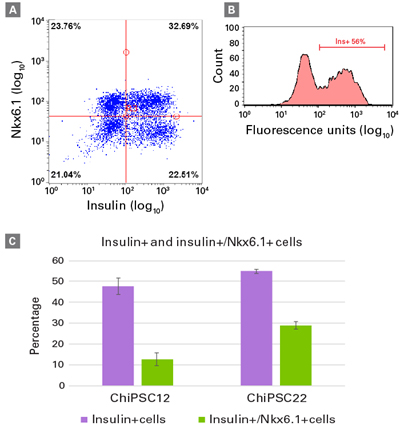
그림 5. Insulin과 Nkx6.1을 동시에 발현하는 hiPSC 유래 beta cell의 정량 분석
Staining for insulin and Nkx6.1 was performed on beta cells from C12 or C22 on Day 14 post-thawing and analyzed by flow cytometry (FC). FC plots are shown for the analysis of one batch of beta cells from C22, with ~33% insulin/Nkx6.1 double-positive cells (Panel A) and ~56% insulin-positive cells (Panels A and B). Percentages of cells expressing insulin and coexpressing insulin/Nkx6.1 are shown for beta cells derived from C12 and C22, respectively (Panel C). Data are presented as mean values ± SEM (n = 3 batches from each cell line).
[원문] Cellartis human iPS cell-derived beta cells for diabetes drug discovery and disease modeling
[참고문헌]
- Ahren, B. Islet G protein-coupled receptors as potential targets for treatment of type 2 diabetes. Nat. Rev. Drug Discov. 8, 369-385 (2009).
- Ashby, J. P. & Frier, B. M. Circulating C-peptide: measurement and clinical application. In Ann. Clin. Biochem (Vol. 18) (1981).
- Bolognini, D. et al. The Pharmacology and Function of Receptors for Short-Chain Fatty Acids. Mol. Pharmacol. 89, 388-398 (2016).
- Gao, T. et al. Pdx1 Maintains β Cell Identity and Function by Repressing an α Cell Program. Cell Metab. 19, 259-271 (2014).
- Horwitz, D. L. et al. Proinsulin, insulin, and C-peptide concentrations in human portal and peripheral blood. J. Clin. Invest. 55, 1278-1283 (1975).
- Layden, B. T., Durai, V. & Lowe, W. L. G-Protein-Coupled Receptors, Pancreatic Islets, and Diabetes. Nat. Educ. 3, 13 (2010).
- Liao, Y., Smyth, G. K. & Shi, W. The Subread aligner: fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 41, e108-e108 (2013).
- Ma, S. et al. β Cell Replacement after Gene Editing of a Neonatal Diabetes-Causing Mutation at the Insulin Locus. Stem Cell Rep. 11, 1407-1415 (2018)
- Marron, M. P. et al. Functional evidence for the mediation of diabetogenic T cell responses by HLA-A2.1 MHC class I molecules through transgenic expression in NOD mice. Proc. Natl. Acad. Sci. U. S. A. 99, 13753-8 (2002).
- Pagliuca, F. W. et al. Generation of Functional Human Pancreatic β Cells In Vitro. Cell 159, 428-439 (2014).
- Pertea, M. et al. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 33, 290-295 (2015).
- Priyadarshini, M. et al. An Acetate-Specific GPCR, FFAR2, Regulates Insulin Secretion. Mol. Endocrinol. 29, 1055-66 (2015).
- Rezania, A. et al. Reversal of diabetes with insulin-producing cells derived in vitro from human pluripotent stem cells. Nat. Biotechnol. 32, 1121-1133 (2014).
- Taylor, B. L., Liu, F. F. & Sander, M. Nkx6.1 is essential for maintaining the functional state of pancreatic beta cells. Cell Rep. 4, 1262-75 (2013).
- Veprik, A. et al. GPR41 modulates insulin secretion and gene expression in pancreatic β-cells and modifies metabolic homeostasis in fed and fasting states. FASEB J. 30, 3860-3869 (2016).
- Yaluri, N. et al. Simvastatin Impairs Insulin Secretion by Multiple Mechanisms in MIN6 Cells. PLoS One 10, e0142902 (2015).
다카라바이오는 당뇨병 및 대사 증후군 연구 모델과 신약 개발을 위한 hiPSC 유래 beta cell의 적용성을 확인하였다.
Introduction
당뇨나 다른 대사 증후군의 연구자에게 정확하고 재현 가능한 질병 모델의 제작은 어려움으로 남아 있다. 현재까지는 인슐린을 생산하는 beta cell을 포함하는 호르몬 생산 세포 군집인 췌장의 랑게르한스섬 (primary pancreatic islets of Langerhans)을 gold standard로 사용되고 있다. 다만 이 Primary cell은 샘플을 확보하기가 어렵고 비용이 많이 소요될 뿐 아니라, 유래한 donor의 유전적 차이로 인해 실험 결과에 변동이 생길 수 있다. 이를 대체하기 위해 단일 기증자에서 유래한 hiPSC로부터 분화한 beta cell이 신약 개발을 위한 효과적인 스크리닝과 insulin 생산 및 분비에 대한 생리학적 실험 모델로 사용될 수 있다.
일관적이고 신뢰도 높은 Beta cell 공급을 위해, 다카라바이오는 췌장 발달 과정을 모방하는 배양 조건을 최적화한 4 step의 표준 분화 프로토콜을 개발하였다 (그림 1). hiPSC는 먼저 내배엽 세포 (definitive endoderm cell)로 분화되고, 이후 pancreatic endoderm, endocrine progenitor cell 단계를 거쳐 최종적으로 성숙한 beta cell로 분화가 완료된다.
그림 1. hiPSC의 beta cell 분화 과정
Our differentiation strategy is based on a four-step protocol that resembles normal embryonic development of beta cells. hiPS cells are first differentiated into Sox17-positive definitive endoderm. Following endodermal commitment, the cells are further differentiated into Pdx1/Nkx6.1 double-positive pancreatic endoderm. Next, differentiation continues into endocrine progenitor cells, which are Ngn3/Nkx2.2 double-positive. Finally, the progenitors mature into terminally differentiated beta cells that are double positive for C-peptide and MafA. Abbreviations: Sox17 = SRY (sex determining region Y)-box 17; Pdx1 = pancreatic and duodenal homeobox 1; Nkx6.1 = NK6 homeobox 1; Ngn3 = Neurogenin 3; Nkx2.2 = NK2 homeobox 2; C-peptide = Connecting peptide; and MafA = MAF BZIP Transcription Factor A.
다카라바이오는 서로 다른 Donor로부터 얻은 hiPSC (ChiPSC12, ChiPSC22)로부터 분화된 총 두 종의 hiPSC 유래 beta cell을 판매하고 있으며, 제품은 세포와 coating substrate, basal media, supplement를 포함한 all-in-one 형태로 제공된다.
지속적인 생산이 가능한 Cellartis® beta cell은 성숙한 beta cell에서 나타나는 특징을 가질 뿐만 아니라, lot에 따른 차이를 최소화 하였고, 개인 간의 차이를 반영하는 기능 평가가 가능하기에 실험에 높은 신뢰성을 부여한다. 당뇨병 연구자들은 Donor HLA type에 따른 변이를 연구할 수 있으며, 역학 연구에 따르면 HLA-A*02:01을 포함하는 class Ⅰ 대립 유전자가 1형 당뇨병에 대한 감수성을 증가시킬 수 있다고 알려져 있다 (Marron et al. 2002). C12와 C22의 HLA type은 표 1에서 확인할 수 있다.
Cellartis hiPS Beta Cells (from ChiPSC12) |
Cellartis hiPS Beta Cells (from ChiPSC22) |
HLA-A*01:01 |
HLA-A*02:01 |
HLA-B*08:01, HLA-B*37:01 |
HLA-B*07:02, HLA-B*40:01 |
HLA-C*06:02, HLA-C*07:01 |
HLA-C*03:04, HLA-C*07:02 |
HLA-DRB1*03:01, HLA-DRB1*11:04 |
HLA-DRB1*13:02, HLA-DRB1*14:01 |
HLA-DQB1*02:01, HLA-DQB1*03:01 |
HLA-DQB1*05:03, HLA-DQB1*06:04 |
HLA-DPB1*01:01, HLA-DPB1*04:01 |
HLA-DPB1*03:01, HLA-DPB1*04:01 |
표 1. ChiPSC12와 ChiPSC22의 donor 및 HLA type 정보
The beta cells originate from donors with different HLA types. ChiPSC12 was sourced from skin fibroblasts from a healthy 24-year-old European/North African male volunteer (76 kg/177 cm). ChiPSC22 was sourced from skin fibroblasts from a healthy 32-year-old European/North African male volunteer (74 kg/179 cm) with a diabetes-susceptible HLA type, HLA-A*02:01.
hiPS cell-derived beta cells express beta-cell-specific genes
다카라바이오는 성숙한 Beta cell에서 존재하는 것으로 알려진 mRNA 전사체가 hiPSC 유래 beta cell에서 발현하는 지 먼저 분석하였다. hiPSC 유래 beta cell에서 모든 전사체가 확인되었으며, 발현 수준을 primary 랑게르한스섬 (islet of Langerhans)과 비교하였다 (그림 2). 특히, 각 Donor의 hiPSC 유래 beta cell의 모든 lot에서 유사한 수준의 mRNA 발현을 확인하였고, lot 간의 높은 재현성을 입증하였다.
그림 2. hiPSC 유래 beta cell에서의 주요 mRNA 발현
mRNA expression analysis, as quantified by RNA-seq, was performed on Cellartis beta cells from C12 and C22 on Day 14 post-thawing (n = 3 batches per cell line) and on primary islets (n = 5 donors). Data are presented as the mean values ± SEM.
*PDX1 = pancreatic and duodenal homeobox 1; GCK = glucokinase; SLC2A1, SLC2A3 = Solute Carrier Family 2 Member 1 and 3; MAFB = MAF BZIP Transcription Factor B; NKX2.2 = NK2 homeobox 2; NKX6.1 = NK6 homeobox 1; and NEUROD1 = Neurogenic differentiation 1.
hiPS cell-derived beta cells synthesize insulin
Beta cell의 주요 기능은 insulin을 합성하고 분비하는 것으로, insulin 합성 과정을 살펴보면 proinsulin이 C-peptide 분자 하나와 insulin 분자 하나, 즉 1:1로 나뉘어 진다 (Ashby & Frier 1981; Horwitz et al. 1975). 따라서, C-peptide와 insulin의 동시 발현이 insulin 생산 능력의 평가 지표로 평가될 수 있다. 그림 3에서 C12와 C22 유래 beta cell 모두에서 C-peptide와 인슐린의 발현이 높은 상관관계를 보임으로써, 세포가 insulin 생산 능력을 갖추었다는 것을 확인할 수 있다.
그림 3. hiPSC 유래 beta cell에서의 insulin 합성
Staining for C-peptide and insulin was performed by immunofluorescence on Cellartis beta cells at Day 14 post-thawing. Colocalization of insulin and C-peptide indicates intracellular insulin synthesis. Scale bar = 100 ㎛.
hiPS cell-derived beta cells express key markers of mature beta cells
몇몇 전사 인자들은 Beta cell의 발달, 증식, 기능에 중요한 역할을 하며, 성숙한 beta cell은 Pdx1, Nkx6.1, 인슐린을 발현한다. Pdx1은 beta cell의 수명을 조절하는 역할로, beta cell이 발현하는 유전자를 활성화 하고 alpha cell에서 발현하는 유전자를 억제한다 (Gao et al. 2014). 반면 Nkx6.1이 결핍된 세포의 경우 delta cell의 분자적 특성을 보인다 (Taylor, Liu, and Sander 2013).
MafA는 성숙한 beta cell marker로써, 분화 과정에서는 발현되지 않아 다른 전사 인자와 확연히 구분된다. 또한 Insulin과 Nkx6.1이 동시 발현하는 것으로 성숙한 hiPSC 유래 beta cell를 확인할 수 있다 (Ma et al. 2018; Pagliuca et al. 2014; Rezania et al. 2014). 그림 4에서는 Cellartis® beta cell에서 insulin과 C-peptide와 함께 주요 beta cell marker인 Nkx6.1, Pdx1, MafA가 발현됨을 확인할 수 있다.
그림 4. hiPSC 유래 beta cell에서 확인한 key beta cell protein의 동시 발현
Staining for insulin and Nkx6.1 (Panel A), insulin and Pdx1 (Panel B), and C-peptide and MafA (Panel C) was performed on beta cells on Day 14 post-thawing. Scale bar = 100 ㎛.
뿐만 아니라, flow cytometry을 이용하여 insulin과 Nkx6.1를 동시 발현하는 세포의 정량 분석 결과를 그림 5에서 확인할 수 있다. 특히 insulin과 Nkx6.1을 동시에 발현하는 세포의 경우, C12과 C22에서 그 비율이 다르게 나타났으며, 이를 통해 HLA type에 따라 일부 유전자 발현에 차이를 확인할 수 있다.
그림 5. Insulin과 Nkx6.1을 동시에 발현하는 hiPSC 유래 beta cell의 정량 분석
Staining for insulin and Nkx6.1 was performed on beta cells from C12 or C22 on Day 14 post-thawing and analyzed by flow cytometry (FC). FC plots are shown for the analysis of one batch of beta cells from C22, with ~33% insulin/Nkx6.1 double-positive cells (Panel A) and ~56% insulin-positive cells (Panels A and B). Percentages of cells expressing insulin and coexpressing insulin/Nkx6.1 are shown for beta cells derived from C12 and C22, respectively (Panel C). Data are presented as mean values ± SEM (n = 3 batches from each cell line).
Code |
제품명 |
용량 |
Y10100 |
1 Kit |
|
Y10106 |
1 Kit |
[원문] Cellartis human iPS cell-derived beta cells for diabetes drug discovery and disease modeling
[참고문헌]
- Ahren, B. Islet G protein-coupled receptors as potential targets for treatment of type 2 diabetes. Nat. Rev. Drug Discov. 8, 369-385 (2009).
- Ashby, J. P. & Frier, B. M. Circulating C-peptide: measurement and clinical application. In Ann. Clin. Biochem (Vol. 18) (1981).
- Bolognini, D. et al. The Pharmacology and Function of Receptors for Short-Chain Fatty Acids. Mol. Pharmacol. 89, 388-398 (2016).
- Gao, T. et al. Pdx1 Maintains β Cell Identity and Function by Repressing an α Cell Program. Cell Metab. 19, 259-271 (2014).
- Horwitz, D. L. et al. Proinsulin, insulin, and C-peptide concentrations in human portal and peripheral blood. J. Clin. Invest. 55, 1278-1283 (1975).
- Layden, B. T., Durai, V. & Lowe, W. L. G-Protein-Coupled Receptors, Pancreatic Islets, and Diabetes. Nat. Educ. 3, 13 (2010).
- Liao, Y., Smyth, G. K. & Shi, W. The Subread aligner: fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 41, e108-e108 (2013).
- Ma, S. et al. β Cell Replacement after Gene Editing of a Neonatal Diabetes-Causing Mutation at the Insulin Locus. Stem Cell Rep. 11, 1407-1415 (2018)
- Marron, M. P. et al. Functional evidence for the mediation of diabetogenic T cell responses by HLA-A2.1 MHC class I molecules through transgenic expression in NOD mice. Proc. Natl. Acad. Sci. U. S. A. 99, 13753-8 (2002).
- Pagliuca, F. W. et al. Generation of Functional Human Pancreatic β Cells In Vitro. Cell 159, 428-439 (2014).
- Pertea, M. et al. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 33, 290-295 (2015).
- Priyadarshini, M. et al. An Acetate-Specific GPCR, FFAR2, Regulates Insulin Secretion. Mol. Endocrinol. 29, 1055-66 (2015).
- Rezania, A. et al. Reversal of diabetes with insulin-producing cells derived in vitro from human pluripotent stem cells. Nat. Biotechnol. 32, 1121-1133 (2014).
- Taylor, B. L., Liu, F. F. & Sander, M. Nkx6.1 is essential for maintaining the functional state of pancreatic beta cells. Cell Rep. 4, 1262-75 (2013).
- Veprik, A. et al. GPR41 modulates insulin secretion and gene expression in pancreatic β-cells and modifies metabolic homeostasis in fed and fasting states. FASEB J. 30, 3860-3869 (2016).
- Yaluri, N. et al. Simvastatin Impairs Insulin Secretion by Multiple Mechanisms in MIN6 Cells. PLoS One 10, e0142902 (2015).
